
Kas dokusu kasılabilir proteinler
içeren farklılaşmış hücrelerden oluşur. Bu proteinlerin biyolojik yapısı belli
başlı organlarda ve bir bütün olarak vücutta, hareketi sağlayacak hücresel
kasılma için gerekli olan güçleri oluşturur. Kas hücrelerinin çoğu mezodermden
kökenlenir ve miyofibriler proteinlerin senteziyle eş zamanlı olarak
gerçekleşen aşamalı bir uzama süreciyle farklılaşır.
Morfolojik ve fonksiyonel
özelliklerine göre memelilerde üç tip kas dokusu tanımlanabilir (Şekil 10-1).
Kas dokusunun her bir tipi onun fizyolojik görevine uygun bir yapıya sahiptir.
İskelet
kası enine
çizgilerime gösteren çok çekirdekli, silindirik, çok uzun demetlerden oluşur.
Kasılmaları, lıızlı, kuvvetli ve genellikle istemlidir. Buna, molekiiler yapıların
birbirleri üzerinde kaymalarına olanak veren, kalın mi- yozin filamanları ve
ince aletin filamanlarının karşılıklı hareketi neden olur.
Kayma için gerekli
olan güçler aklini miyo- zine bağlayan köprülerdeki zayıf etkileşimlerle
oluşturulur. Kalp kası da enine çizgilenmeler gösterir ve birbirine paralel
uzanan dallanmış özel hücrelerden meydana gelir. Uç uca geldikleri bölgelerde,
sadece kalp kasında bulunan yapılar olan interkalar
disklerle birbirlerine
bağlanırlar. Kalp kası istemsiz, güçlü ve ritmik kasılır. Düz kas, ışık mikroskobunda
çizgilerime göstermeyen iğ biçimli hücre gruplarından meydana gelir, bunların
kasılması yavaştır ve istem dışı çalışır.
Kas hücrelerinin bazı organelleri
diğer hücrelerdeki karşılıklarından farklı isimlere sahiptir. Kas hücre
sitoplazması (kas lifçikleri dışında) sarkoplazma, (Yun. setrkos, et + plcts- »ıcı, nesne), düz endoplazına retikulumu sarkoplazma re-
tikulumu olarak
adlandırılır. Sarkolemma (Yun. sarkos + lenıma, örtü) hücre zan ya da plazmalemmadır.
İskelet kası demetler halinde uzayan,
çok uzun (30 cm’nin üzerinde) silindirik, 10-100 ////»çapında çok çekirdekli
hücrelerin oluşturduğu kas liflerinden oluşur. Çok çekirdeklilik, embriyonik
tek çekirdekli miyoblastların (kas hücresi öncülü) birleşmesi sonucunda
meydana gelir. Oval çekirdekler genellikle periferde, hücre zarının hemen
altında bulunur. Bu tipik çekirdek yerleşimi iskelet kasını, çekirdekleri merkezde
yer alan kalp ve düz kastan ayırmaya yarar.
İskelet kası liflerinin
çaplarındaki çeşitlilik yaş, cinsiyet, beslenme durumu ve bireyin fiziksel eğitimine
bağlıdır. Egzersizin kas yapısını geliştirdiği ve yağ depolarını azalttığı
konusunda ortak görüş bulunmaktadır. Kasta bu artma yeni miyo- kas
lifçiklerinin oluşumu ve her bir kas lifinin çapının artmasına bağlıdır.
Özelliğini hücre hacminin artışından alan bu olaya hipertrofi denir (Yun.
hyper,
aşırı + trophe, beslenme). Hücre sayısındaki artışa bağlı
doku büyümesine hiperpla- zi denir. (Hyper, + Yun. plasis, şekillenme). Hi- perplazi, mitoz bölünme
özelliğini kaybetmemiş olan düz kasta görülmesine karşın, gerek iskelet
kasında, gerekse kalp kasında izlenmez. Hem hi- perplazi, hem de hipertrofi,
gebelik sırasında ute- rusta olduğu gibi birçok organda sık ortaya çıkar.
Değişik tiplerdeki kasları oluşturan
kas lifi kütleleri gelişigüzel değil, düzenli demetler halinde dizilirler ve epimisyum (Yun. epi, üst + ınys, kas) adı verilen tıkız bağ dokusu
bir kılıf ile dıştan tümüyle sarılırlar. (Şekil 10-2, 10-3 ve 10-4). Epi-
misyumdan içeriye doğru ince bağ dokusu bölmeleri uzanarak bir kasın içindeki
lif demetlerini sarar. Kas liflerinden oluşan her bir demetin etrafındaki bağ
dokusuna perimis- yum (Yun. peri, çevre + ınys, kas) denir. Her kas lifi ise bazal lamina ve retikiiler
liflerden oluşmuş ince bir bağ dokusu tabakası olan endomisyum (Yun. enclotı içinde, + mys) ile sarılıdır.
Bağ dokusunun en önemli ödevlerinden
biri, kasılan kas hücrelerinin oluşturduğu güçleri mekanik olarak iletmektir.
Çünkü çoğu kez kas hücreleri tek başına kasın bir ucundan diğer ucuna uzanmaz.
Kan damarları kasa bağ dokusunun
oluşturduğu bölmeler içinde girer ve kas lifleri arasında, onlara koşut olarak
zengin bir kılcal damar ağı oluşturur. Kılcal damarların endoteli kesintisiz
tiptedir ve bağ dokusunda lenfatikler de bulunur.
Bazı kaslar kas tendon bileşkesinin
oluştuğu uçlarda ortadan kalkar. Elektron mikroskobik bulgulara göre bu geçiş

Şekil
10-2. İskelet
kasının yapısı ve işlevi. Sağdaki çizimde geniş bir segmentte kasın ayrıntıları
görünmektedir. Renkli çizgiler endomisyum, perimisyum ve epimisyumdur.

Şekil 10-3. İskelet kasının enine kesitinde
tip I ve tip III kolajen ile hücre çekirdekleri görülmektedir. Endomisyum ok
başlarıyla, perimisyum oklarla gösterilmektedir. Solda epimis- yumun bir bölümü
yer almaktadır. Pikrosirius-hematoksilen boyası. Yüksek büyütme.

Şekil 10-4. İskelet kasının endomisyumda
bulunan bir protein bileşeni olan laminine yönelik immünhistokimya boyası
yapılmış kesiti. Laminin kahverengi gölgeler halinde izlenmektedir. Sağ üst
köşede küçük bir sinirin hafif çapraz kesiti bulunmaktadır. Sinir liflerinin
çevresinde de laminin bulunur.

|
||||||||||||||||||||

|
||||
bölgesinde tendonun kolajen
liflerinin, kas liflerinin plazma- lemmasmdaki karmaşık kadanaların içine
gömüldükleri görülür.
Işık mikroskobunda gözlendiği gibi,
uzamına kesiti alınan kas lifleri açık ve koyu bantlar halinde ardışık
sıralanan enine çizgilenmeler gösterir (Şekil 10-6’dan 10-9’a). Koyu olarak
gözlenen bantlar A bandı (anizotrop) olarak isimlendirir; açık gözüken bantlar ise I bantları
(izotrop, yani polarize ışık
altında değişmeyen) olarak adlandırılır. Elektron mikroskopta her I bandının Z çizgisi denilen enine koyu bir çizgiyle ikiye
ayrıldığı gözlenir. Kasılabilir yapının yinelenen en küçük parçası olan sarkomer (Yun. sarkos + mere, bölüm) bir Z
çizgisinden diğer Z çizgisine uzanan kısımdır (Şekil 10-10 ve 10-11) ve
dinlenme halindeki kasta yaklaşık 2.5 jam. uzunluğundadır.


Şekil 10-9. İskelet kasının uzamına kesiti.
Kas hücrelerindeki çizgîlenme ve orta düzeyde kolajen (sarı) miktarı
görülmektedir. PSP boyası. Büyük büyütme.

Şekil 10-10. İribaş (kurbağa larvası) iskelet
kasının elektron mikroskop fotoğrafı. A, I, H bantları ve Z çizgisiyle sar-
komer yapısı görülmektedir. Sarkomerdeki kalın ve ince filamanlar şeklin alt
bölümünde şematik olarak gösterilmektedir. Burada da görüldüğü gibi, hem
suda, hem karada yaşar (amfibi) canlıların kaslarındaki triadlar her
sarkomerin Z çizgisi hizasında yer almaktadır. Memelilerin kaslarında ise her
sarko- merde A-l bantlarının kesişim hizasında yer alan karşılıklı 2 adet
triad bulunur (bkz. Şekil 10-16). X 35.000. (KR Porter’in
izniyle.)

Kas lifinin sarkoplazması kas lifçiği
adı verilen uzun, si- lindirik, filamanlı demetlerle doludur. 1-2 pm çapında
olan ve kas lifinin uzun eksenine koşut olarak uzanan kas lifçik- leri ucuca
zincir biçiminde düzenlenmiş sarkomerlerden oluşur (Şekil 10-10 ve 10-11).
Bitişik kas lifçiklerindeki sar- komerlerin kenarlara yerleşik tutunma düzeni
tüm kas lifinin enine çizgilenmeler gösteren karakteristik bir yapı göstermesine
neden olur.

Elektron mikroskobik çalışmalar
aslında sarkomerin bu düzeninin kalın ve ince olmak üzere 2 tip filamanın, kas
lifçiklerinin uzun eksenine koşut olarak simetrik dizilimiyle meydana
geldiğini göstermiştir (Şekil 10-10 ve 10-11). Kalın filamanların boyu 1,6
pırı, eni 15 nm’dir; sarkomerin orta bölümünde, A bandını oluştururlar. İnce
filamanlar, kalın fila- manlar arasında bir uçları Z çizgisine tutunmuş
şekilde, onlara koşut olarak uzanır. (Şekil 10-10 ve 10-11). İnce filaman-
ların boyu 1,0 pm, eni 8 nm’dir. Bu düzenleniş sonucunda I bandları, ince
filamanların kalın filamanların üzerine gelmeyen kısımlarından meydana gelir.
A bandı ise, esas olarak kalın ve ince filamanlarla kalın filamanların üst üste
gelen kısımlarından oluşur. A bandının ortasında II bandı olarak isimlendirilen
daha açık bir alan vardır. Burası sadece miyo- zin molekülünün çubuk benzeri
kısımlarından oluşmuş bir alana karşılık gelir. M bandı M çizgisiyle ikiye
ayrılır. Burası komşu kalın filamanlar arasında yan bağlantıların bulunduğu
bölgedir. (Şekil 10-11). M çizgisinin esas proteini kreatin kinaz’dır. Kreatin
kinaz, bir fosfat grubunun fosfokreatinden (fosfat grupları şeklindeki yüksek
enerji deposu) ADP’ye aktarımında kolaylaştırıcı görevi görür. Böylece kasın
kasılması için gerekli ATI3 sağlanır.

İnce ve kalın filamanlar A bandında
üst üsle binerler. Bu bölgenin enine kesitinde, her kalın filamanı bir altıgen
oluşturacak şekilde 6 ince filaman kuşatır (Şekil 10-10 ve 10-11).
Çizgili kas fikımanları birkaç
protein içerir; başlıca -i protein aktin, tropomyozin, troponin ve miyozindir.
İlk üçü ince fikımanları oluştururken, miyozin kalın filamanı yapar. Aktin ve
miyozin çizgili kasın toplam proteininin %55’ini oluşturur.
Aktin çift sarmal şekilde birbiri
etrafına dolanmış 5,6 nm çapında 2 sıra globiiler monomerden (G aktin) oluşan
uzun filaman oluşturan polimerlerden (F aktin) ibarettir (Şekil 10- 11).
G-Aktin moleküllerinin önemli bir özelliği de yapısal asimetri göstermesidir.
G-Aktin molekülleri F-aktini oluşturmak üzere polimerize olduğunda, belirgin
kutupsallık gösteren filamanlar oluşturarak arkadan öne bükülürler (Şekil
9-
13).
Her bir G-aktin monomeri miyozin için bağlanma bölgesi içerir (Şekil 10-1-i). Z
çizgisi üzerinde dik açıyla tutunan aktin filamanlar! çizginin her iki
tarafında karşıt ku-
Şekil 10-13. Şematik çizimde, ince filamanın 3
ana proteini olan aktin, tropomiyozin ve troponinin üç boyutlu düzlemdeki
ilişkisi görülmektedir. Şeklin üst kısmında bileşenler tek, alt kısmında ise
zincirleşmiş şekilde izlenmektedir. Globüler aktin molekülleri tek bir yöne
doğru kutuplaşır ve zincirleşir. Her tropomiyozin molekülü yedi tane aktin molekülünün
üzerinde uzanır. Tnl, TnC, TnT troponinin alt birimleridir.

Şekil 10-14. Troponinin TnC birimine Ca+2’un
bağlanmasıyla aktin üzerindeki miyozin bağlayıcı bölüm açığa çıkar ve kas
kasılması başlar. İkinci basamakta, miyozin başı aktine bağlanır ve ATP ADP’ye
parçalanır, üretilen enerji miyozin başının hareketini sağlar. Miyozindeki bu
değişiklik sonucu ince filaman bantları kalın filaman üzerinde kayar. Tek bir
kasılma sırasında birçok kez yinelenen bu olay aktin ve miyozinin üst üste binmesini
sağlar ve tüm kas lifinin kısalmasıyla sonuçlanır (İzin alınarak basım, Ganong WF: Review of Medical Physiology, 14th ed. Appleton & Lange, 1989).
tupsallık gösterir (Şekil 10-11). Z
çizgisinin ana bileşeni olan a-aktinin proteininin, aklin filamanlarına bu
bölgede tutunduğu düşünülmektedir. a-Aktinin ve desminin (bir ara Ulaman proteini)
komşu sarkomerleri birbirine bağladığına ve bu şekilde bu kas 1 ildiklerini
belli bir düzen içinde tuttuğuna inanılır.
Tropomiyozin 2 polipeptid zincir içeren -İ0 nm
uzunluğunda ince uzun bir moleküldür. Hu moleküller birbirine dolanmış iki
aklin uzantısı arasındaki oluğun dıs kenarlan boyunca aktinin alt birimleri
üzerinde filamankır oluşturarak baştan sona kadar uzanan moleküllerdir (Şekil
10-13).
Troponİn, tropomiyozine giiclü bir şekilde
tutunan TnT, kalsiyum
iyonlarını bağlayan TnC ve aktin-miyozin iliskisini bozan Tnl olmak üzere 3 alt birimden oluşan bir
komplekstir. Bir troponin kompleksi her tropomyozin molekülü üzerinde belirli
bir noktaya tutunur (Sekil 10-13).
İnce filamanlarda, her bir
tropomiyozin molekülü yedi tane G-Akıin molekülüne uzanır ve bunun yüzeyine bir
troponin kompleksi tutunur (Şekil 10-13).
Miyozin çok daha büyük bir komplekstir.
(MA-500.000). Miyozin, birbirine eş iki ağır zincir ve İki cilt hafif zincire
ay- rıstırıhıbilir. Miyozinin ağır zincirleri birbirine sarmalanmış iki ağır
zincirden meydana gelen ince, çubuğa benzer moleküllerdir (150 nm uzunluğunda
2-3 nm kalınlığında). Her ağır zincirin bir ucundaki küçük globüler uzantılar.
ATP bağlamanın yanışını ATP’yi hidrolize edecek enzimatik kapasiteye sahip
basları oluşturur ve aklin bağlama özelliği sergiler. Hu ı hafif zincir bas ile
ilişkilidir (Sekil 10-11). Her bir kalın Ulamandaki yüzlerce miyozin molekülü
çomağa benzer kısımları üst iiste gelecek ve globüler basları bir diğerinin
ucuna yönelecek şekilde düzenleniş gösterir (Şekil 10-11).
Çizgili kasın ince kesitlerinde kas lifleri ince ve kalın
fi- [amanlar arasında karşılıklı köprülerin bulunduğu görülür. Bu köprüler
miyozin molekülünün baş kısmından ve çubuğa benzeyen parçasının kısa bir
bölümünden meydana gelir. Bu köprüler kimyasal enerjiyi doğrudan mekanik
enerjiye çevirmede rol üstlenir (Sekil 10-1 -i).
Sarkoplazma retikulum zarının
depokırizasyonu kas hücresinin yüzeyindeki özelleşmiş kas-sinir bağlantısında
başlar ve Ca+- iyonlarının salıverilmesi ile sonuçlanır. İçerideki
sarkoplazma retikulumu sarnıçlarından Ca+~'un salıverilmesini
etkilemek için depokırizasyon sinyallerinin tüm hücre boyunca dalga halinde
ilerlemesi gerekir. Periferik kas lifçikle- ri merkezde yerleşik kas
lileiklerinden daha önce kasılır. Düzenli bir kasılmanın sağlanması için,
iskelet kası enine (T) tiibüllere sahiptir (Şekil 10-15). Sarkolemmanın
parmağa benzeyen bu girintileri kompleks bir tübül ağı meydana getirir ve kas
lifçikleri içindeki her sarkomerin A-l bantlarının sınırlarını kuşatır (Sekil
10-16 ve 10-17).
Sarkoplazma retikulumunun T tübüle
komşu karşılıklı iki kenarı genişleyerek terminal sarnıçları yaparlar. 'I’
tübül ile birlikte iki yandaki sarkoplazma retikulumu bölümünden oluşan bu
özelleşmiş komplekse triad denir (Sekil 10-10, 10- 16, 10-17). Triadda sarkolemmanın
oluşturduğu T tıibüllerin depokırizasyonu, sarkoplazma retikulumu membramna aktarılır.
Yukarıda anlatıldığı gibi ortamda Ca-+
iyonları varsa kas kasılır, bulunmadığında gevşer. Sarkoplazma retikulumu hızlı
kasılına ve gevşeme döngüleri için gerekli olan kalsiyum akı-
|
|||||||

Şekil 10-16. Maymun iskelet kasının enine
kesitinin elektron mikroskop fotoğrafı. Komşu kas lifçikleri arasında
mitokondriyumlar (M) görülmektedir. Ok başları bu kasta, her bir sarkomer için
2 tane olmak üzere, A-l bantlarının iki yanlı birleşim hizasında bulunan
triadları göstermektedir. A, A bandı; I, I bandı; Z, Z çizgisi x 40,000. (Alıntı,
Junqueira LCU, Salles LMM’nin izni ile Ultra-Estruture e Funçao Cel- lular. Edgard
Blücher, 1975.)
sim düzenler. Sarkoplazma reıikulumu .sistemi her bir kas
lif- çiğini kuşatan düz endoplazma reıikulumu sarnıçlarının meydana getirdiği
dallı bir ağdan ibarettir. Sarkoplazma retikulu- nuı zarının sinirler aracılığı
ile depolarizasyonunun ardından, sarkoplazma reıikulumu içinde yoğunlaşmış Ca~+
iyonları kalın ve ince filamanların üsl üste bulunduğu bölüme pasif olarak
bırakılır ve burada troponine bağlanıp aklin ve miyozin arasında köpriilesmeye
neden olur. Hücre zarı depolarizasyo- nu sona erdiğinde, sarkoplazma reıikulumu
Ca-+ deposu olarak davranarak Ca+~ u aktif transponía sarnıçlara
geri gönderir ve bu da kasılma işleminin sona ermesiyle sonuçlanır.
Dinlenme halindeki sarkomerler kısmen
üst üste binmiş kalın ve ince filamanlardan oluşmaktadır. Kasılma sırasında
hem ince hem de kalın filamanlar kendine özgü boylarını korurlar. Kasılma bu
filamanlardaki kısalmaya bağlı olmadığı için filamanların üsl üste binme
ölçüsünün artmasına bağlanabilir. Kas kasılmasında kayan filaman hipotezi
yaygın olarak benimsenmiştir.
Aşağıda, kasılma döngüsü sırasında
aklin ve miyozin fi- lamankırı arasındaki etkileşim kısaca tanımlanmaktadır.
Dinlenme sırasında ATI1 miyozin başındaki ATP-az kısmına bağlanır.
Ancak hidroliz hızı çok yavaştır. Miyozinin ATP'yi hızlı olarak parçalayarak
enerjinin ortaya çıkmasını sağlamak için bir kofaktör olarak aklinin varlığına
gereksinimi vardır. Dinlenme durumundaki kasla miyozin aklin ile bağlantı kuramaz,
çünkü aktin molekülleri üzerinde miyozin başlarının bağlanması için uygun olan
kısımlar F-aktin filamanı üzerindeki iroponin-tropomiyozin kompleksleriyle
örtülmüş durumdadır (Şekil 10-l-ı üstte). Cai+ iyon derişimi
yeterince yüksek olduğunda bunlar troponinin TnC alt birimine bağlanırlar. Üc
troponin alt biriminin konumları değişir ve tro- pomiyozin molekülünü aktin
sarmalı oluğunun İçine doğru iter (Sekil 10-14). Bu, globüler aktin bileşenleri
üzerindeki miyozin bağlayan bölgeyi açığa çıkarır, böylece aktin, miyozin
molekülünün başı ile birleşmek üzere serbest kalır.
Kalsiyum iyonlarının TnC birimine
bağlanması, miyozin- ATP’nin aktif komplekse dönüştüğü döneme karşılık gelir.
Miyozin bası ve ince Ulamanın G-aktin alt birimi arasındaki bağlanmanın sonucu
olarak ATP, ADP ve Pi'ye dönüşerek enerji açığa çıkar. Bu akıivite miyozin
başının ve çubuk benzeri bölümün bir parçasının (bükülme bölgesi) bükülerek
biçim değiştirmesine sebep olur (Şekil 10-14). Aktiniti miyo- zine bağlı olması
nedeniyle miyozin basının harekeli aklini miyozin üzerinde çeker. Sonuç olarak
ince filaman A bandı içine ileri itilir.
Çok sayıda miyozin basının kalın
Ulamandan uzanmasına karşın kasılma anında az sayıda bas, mevcut aklin bağlayan
kısımlarla karsı karşıya gelir. Bağlı miyozin başları aklini hareket ettirirken,
yeni aktin-miyozin bağlantılarının karsı karsıya gelmesini de sağlar. Eski
aktin-miyozin bağlantıları, yalnızca miyozin yeni bir ATP molekülü bağladıktan
sonra çözülür. Bu işlem aynı zamanda miyozin başını serbestleştirir ve onu
başka bir kasılma döngüsüne hazırlar. Eğer ortamda hiç ATP yoksa, aktin-miyozin
kompleksi durağan hale geçer. Bu durum, ölümden sonra ortaya çıkan asın kas
sertliğine (ölü sertliği - rigor mortis) neden olur. Tek bir kas kasılması
yüzlerce bağlantının kurulma ve koparılma döngüleri sonucu ortaya çıkar. İnce
ve kalın filamanların tam olarak üst üsle binmesiyle oluşan kasılma aktivitesi
Ca~+ iyonlarının iroponinden ayrılmasına ve iroponin-tropomiyozin
karmasının miyozin bağlayan kısımları tekrar örtmesine kadar devam eder.
Kasılma sırasında ince filamanlar A
bandına girdikçe I bandının boyu kısalır. İnce filamanlar kalın filamanlarla
ta-

Şekil 10-17. Memeli iskelet kasının bir
parçası. Kısmen kesiti görünen sarkolemma ve diğer kas lifi elemanları: T
sisteminin girintileri her sarkomerde 2 adet olmak üzere, A ve I bantları
arasındaki geçiş hizasında gözlenir. Sarkoplazma retikulumunun terminal
sarnıçları ile ilişkisi triadları oluşturur. Kas lifçikleri arasında çok
sayıda mitokondri bulunur. Kas lifçiklerinin kesit yüzeyinde ince ve kalın
filamanlar görünmektedir. Sarkolemmayı bir bazal lamina ve retiküler lifler
sarar. (Alıntı, Krstic RV’nin izniyle Ultrastructure of the Mammalian
Celi. Springer Verlag 1979.)
maınen örtüştüğünde H bandının (A bandının yalnızca kalın filamanları içeren kısmı) eni kısalır. Sonuçta her sarkomer ve buna bağlı olarak tüm hücre (lif) büyük oranda kısalır (Şekil 10-18).
Miyelinli motor sinirler perimisyuma
ait bağ dokusu içinde dallanır ve sonlandıkları bölgede çok sayıda ince dal
oluştu- rur. Sinir bağlantısı bölgesindeki sinir, miyelin kılıfını yitirir ve
kas hücresi yüzeyine oturan geniş bir uç oluşturur. Bu yapı motor son plak ya da miyonöral bağlantı
olarak isimlendirilir
(Şekil 10-18). Bu bölgede akson Schwann hücrelerine ait ince bir
sitopkızma tabakası tarafından sarılır. Aksonun ucunda çok sayıda mitokondri
ve sinaps keseleri bulunur. Bu keseler asetilkolin nörotransmiteri (sinir aktargaç-
ları) içerir. Akson ve kas arasında sinaps yarığı denilen bir boşluk vardır. Bu
boşluğun içinde şekilsiz bir bazal lamina matriksi bulunur. Kas sinir
bağlantısında, sarkolemma derine uzanan bağlantı kıvrımları şeklinde katlanmalar yapar.
Sarkolemma katlanmalarının altında çok sayıda çekirdek, mitokondri, ribozomlar
ve glikojen granülleri bulunur.
Motor son plak uyarıldığı zaman akson
terminalinden asetilkolin salıverilir, yarık boyunca dağılır ve bağlantı kıvrımlarının
sarkolemması üzerindeki asetil kolin reseptörlerine tutunur. Transmiterİerin
bağlanması sarkolemmayı sodyuma geçirgen hale getirir. Bu da zar
depolarizasyonu ile
sonuçlanır. Sinaps yarığının bazal laminasına tutunan koii- nesteraz enzimi
asetilkolini hidrolize eder. Transmitterin sar- kolemmadaki reseptörlerle uzun
süre temas etmesinin önlenmesi açısından asetilkolinin parçalanması gerekir.

Motor son plakta başlayan
depolarizasyon kas hücresinin yüzeyi boyunca ve enine tübül sistemi
aracılığıyla lifin derinliklerine doğru yayılır. Her triadda, depolarizasyon
sinyali sarkoplazma retikulumuna geçer ve sonuçta kasılına döngüsünü başlatan
Ca2+ serbestlenmesi ile sonuçlanır. Depolarizasyon sona erdiğinde
Ca-+ aktif olarak sarkoplazma retikulumıı sarnıçlarına geri alınır
ve kas gevşer.
-)
ı
Miyastenia
gravis, sinir kas bağlantısında sar- kolemma içindeki işlevsel yönden aktif
asetilkolin reseptörlerinin sayıca azalmasının neden olduğu, ilerleyici kas
güçsüzlüğü ile kendini gösteren otoimmün bir hastalıktır. Bu azalma dolaşımdaki
antikorların bağlantı kıvamlarındaki asetil kolin reseptörlerine bağlanarak,
normal kas sinir ilişkisini inhibe etmesine bağlıdır. Vücut bu durumu
düzeltmeye kalkıştığında, etkilenmiş reseptörleri içeren zar parçaları hücre
içine alınır, lizozomlar tarafından sindirilir ve yerine yeni reseptörler konur.
Ancak yeni reseptörler de aynı antikorlar tarafından asetil koline yanıt
veremez hale getirilir ve hastalık ilerlemeye devam eder.
Tek bir sinir lifi (akson) bir kas
lifini uyarabilir ya da dallanarak 160 ya da daha fazla sayıda kas lifinin
uyarılmasını sağlayabilir. Birden fazla uyarımın gerçekleştirildiği durumlarda,
bir sinir lifi ve onun uyardığı tüm kaslara motor birim adı verilir. Çizgili
kas lifleri aşamalı kasılmazlar. Ya tümü kasılır ya da hiç kasılmazlar (hep ya
da hiç kuralı). Kasılmanın gücünü değiştirmek için bir kas demeti içindeki liflerin
tümünün aynı anda kasılmamaları gerekir. Kaslar motor birimlere ayrıldıkları
için tek motor aksonun uyarımı bu akson tarafından uyarılan kas liflerinin
sayısıyla orantılı bir gerilme yaratır. Motor birimlerin sayısı ve her birimin
boyutunun değişken olması kasın kasılma gücünü kontrol eder. Bir kasın duyarlı
hareket oluşturma yeteneği, o kasın motor birimlerinin boyutuna bağlıdır.
Örneğin, göz kaslarının iyi kontrol edilmesi gerektiğinden bu kasların her lifi
farklı bir sinirden bağlantı alır. Kol ve bacak kasları gibi daha kaba
hareketleri gerçekleştiren büyük kaslarda, tek bir sinir lifi 100’den fazla kas
lifini uyarır.
İnsandaki bütün çizgili kaslarda kas
iğciği adı verilen kap- süllü proprioseptörler (Lal. propriııs, kendine ait + ccıpio, almak) bulunur (Şekil 10-19). Bu
yapılar içi sıvı dolu bir boşluğu saran bağ dokusu kapsülünden ibarettir. Bu
sıvı dolu alanın içinde az sayıda uzun ve kalın kas lifi ile bir miktar kısa ve
ince lif bulunur. Bunlara topluca intrafüzal lifler denir. Kas iğciğinin içine
birkaç duysal sinir lifi girer ve burada ekstrafüzal kas liflerindeki uzunluk
(gerilme) değişimini tespit ederek bu bilgileri omuriliğe iletir. Omurilik te
değişken karmaşıklıktaki refleksler etkinleşerek vücut durusunu sağlar ve
yürüme gibi motor aktivitelere katılan antago- nist kas gruplarının
aktivitesini düzenler.
Tendonkırda ise kas liflerinin
lendona tutunduğu bölgenin yakınında bağ dokusu kılıfı çok sayıda kalın
kollajen lif demetini sarar. Bu demetler miyotendinöz bağlantıyı oluşturan
kollajen lifler olarak devam eder. Duysal sinirler bağ dokusu kapsülüne girer.
Bu yapılar tendonlardaki gerilme değişikliklerini algılayarak proprioseptör
görevini üstlenir ve bunlara Golgi tendon organları adı verilir (Şekil 10-20).
Bu yapılar gerilme artışına duyarlı
oldukları için, değişik düzeylerde kas gücü gerektiren hareketleri
gerçekleştirmede harcanacak güç miktarını kontrol eder.
İskelet kası hücreleri, kimyasal enerji ile aralıklı olarak
gerçekleştirilen yoğun mekanik ise yüksek derecede uyum gös-

Şekil 10-19. İntrafüzal liflerle (yapısı
değişmiş kas lifleri) si- naps yapan aferent ve eferent sinir liflerinin
görüldüğü kas iğciği. İntrafüzal lifler üzerindeki karmaşık sinir uçları dikkati
çekmektedir. Bu çizimde biri küçük çaplı, diğeri çekirdeklerle dolu,
genişlemiş olmak üzere iki tip intrafüzal lif gösterilmektedir. Kas iğcikleri
bedenin duruşunun sinirsel kontrolünde rol üstlenir ve karşıt etkili kasların
hareketinin eşgüdümlü olmasını sağlar.
|
mı
|
Golgi tendon organı
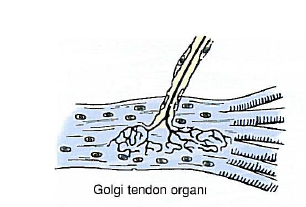
Şekil 10-20. Golgi tendon organının çizimi. Bu
yapı, tendon- lar arasındaki gerginlik farklılıkları hakkında bilgi toplar ve
verileri merkezi sinir sistemine gönderir. Merkezi sinir sistemi bu verileri
işler ve hassas kas kasılmalarını düzenler.
terirler. Bu aktivite dalgaları ile
baş edebilmeleri için enerji depolamaları gerekir. Gerekli enerjinin çoğu, her
ikisi de enerjiden zengin fosfat bileşikleri olan fosfokreatin ve ATP şeklinde
depolanır. Kimyasal enerji aynı zamanda kas ağırlığının yaklaşık % 0,5-1’ini
oluşturan glikojen depolarında mevcuttur. Kas dokusu fosfokreatin ve ATP
şeklinde depolanan enerjiyi, glikoz ve yağ asitlerini yıkarak üretir. Yağ asitleri
mitokondriyum ıııatriksinde bulunan P-oksidasyon enzimleri ile asetata
parçalanır. Asetatın sitrik asit döngüsü yoluyla okside olmasıyla ATP şeklinde
depolanan enerji ortaya çıkar. İskelet kası kısa süreli egzersizde glikozu
(esas olarak kastaki glikojen depolarından elde edilir) hızlı bir şekilde
laktata meta bol ize ederler; bu da toparlanma dönemi sırasında giderilen bir
oksijen açığına neden olur. Bu tür egzersizlerde oluşan laktat, kramplara ya
da İskelet kası ağrılarına neden olur.
Morfolojik, histokimyasal ve işlevsel durumlarına göre iskelet kası lifleri
tip l (yavaş) ve tip II (hızlı) lifler şe
Kasın, kırmızı, beyaz ya da ara lif tiplerine farklılaşmasını
sinir dağılımı kontrol eder. Kırmızı ve beyaz liflerin sinirlerinin kesilerek
çapraz olarak ucuca getirilip İyileşmeye bırakıldığı deneylerde kas lifleri
morfolojik ve fizyolojik durumlarını onları uyaran sinire göre değiştirirler.
Kasın dener- vasyonu (sinirlerini yitirmesi) lif atrofisine ve felce yol açacaktır.
Glikojen, sarkoplazmada kaba granüller şeklinde
bol olarak bulunur (Şekil 10-15). Kas kasılması sırasında hareketi sağlayacak
enerjiyi depolamaya yardımcı olurlar.
Sitoplazmanın başka bir bileşeni de,
bazı kaslarda koyu kırmızı rengin asıl sorumlusu olan hemoglobine benzer oksijen
bağlayan bir protein olan miyoglobindir. Miyoglobin bu tip liflerde yüksek
düzeyde oksidatif fosforilasyon ¡cin gerekli olan oksijeni depolayan bir
pigmenttir. Bu nedenle de derin suda yaşayan memelilerin (örn., fok, balina)
kaslarında büyük miktarlarda miyoglobin bulunur. Aktivitesini uzun süre
korumak zorunda olan kaslar genelde kırmızıdır ve yüksek yoğunlukta miyoglobin
içerir.
Olgun kas hücreleri bu dokularda gözlenen düşük seviyedeki
protein sentezine uygun olarak, az miktarda graniil- lii endoplazma retikulumu
ve ribozomlar barındırır.
.
İskelet kası
Embriyonun gelişimi sırasında ilkel
kalp tüpünün splanknik mezoderm hücreleri zincire benzer sıralar halinde
dizilirler. Kalp hücreleri, iskelet kasındaki birleşik hücreler gibi kaynaşmak
yerine, uzantıları arasında kompleks bağlantılar oluştururlar. Zincirdeki
hücreler çoğunlukla dallanır ve komşu zincirdeki hücrelere tutunurlar. Bu
şekilde kalp, sıkı bir örgü demeti halinde düzenlenmiş olan hücrelerden oluşur,
böylece kalp karıncıkları derinlemesine etkileyen tipik kasılma dalgasının
oluşması sağlanır.
Olgun kalp kası hücrelerinin çapı
yaklaşık 15 pm, boyu ise 85-100 pm’dir. Bunlar iskelet kasında olduğu gibi
enine çizgiler şeklinde bantlaşma gösterir. Çok çekirdekli iskelet kasının
tersine her kalp kası hücresinin yalnız 1 ya da 2 tane, merkezde yerleşmiş,
soluk boyanan çekirdeği vardır. Kas hücrelerini, zengin bir kapiler ağ içeren
endomisyuma ait ince bir bağ dokusu kılıfı sarar.
Kalp kasını diğerlerinden ayıran tek
tipik özellik kalp hücrelerinin oluşturduğu, zincirleri düzensiz aralıklarla kesen
koyu boyanmış enlemesine çizgilerin bulunmasıdır. (Şekil 10-22 ve 10-23).
Bunlar interkalar diskler olarak bilinen, birbirine komşu kalp kası hücreleri
arasındaki bitişik yüzeylerde bulunan oldukça kompleks bağlantılardır (.Şekil
10-
24,
10-25 ve 10-26). Bağlantılar düz bir hat şeklinde ya da basamak biçiminde
gözlenebilir. Basamaksı bağlantılarda 2 bölge ayırt edilebilir; enine bölüm lifleri dik açı ile keserken, yan bölüm miyofilamanlara koşut uzanır.
Disklerde başlıca 3 bağlantı yapısı vardır. Fasya adherens, diskin enine bölümlerindeki hücre
zarı özelleşmeleri olup, terminal sarkomerlerin aletin filamanları için tutunma
bölgesi işlevi görürler. Bunlar aslında, yarı-Z bantlarına karşılık gelir. Ma- küla
adlıerensler (desmozomlar)
sabit kasılma aktivitesi sırasında kalp kası hücrelerini, ayrılmayacak biçimde
birbirlerine bağlarlar. Diskin lateral bölümünde yer alan aralık bağ-
lantılan (neksus)
komşu hücreler arasında iyon bütünlüğünü sağlar (Şekil 10-26). İyon eşleşmesi,
tek tek hücrelerden oluşan bu zincirin bir bütün olarak davranmasını koşullaya-
rak, kasılma sinyalinin hücreden hücreye bir dalga halinde yayılmasını
sağlaması açısından önemlidir.
Aslında, kalp kası hücrelerindeki
kasılma proteinlerinin yapısı ve işlevi, iskelet kasındaki gibidir. Ancak T
tübiil sistemi ve sarkoplazma retikulumu, kalp kası hücrelerinde çok düzenli
değildir. 'I' tübiilleri kalp karıncığı kaslarında iskelet kasındakinden daha
fazla ve daha büyüktür. Kalp kasının T tübiilleri memeli iskelet kasındaki gibi
A-I bağlantısında de-


ğil Z bandı hizasında yer alır.
Sarkoplazma retikulıımu çok iyi gelişmemiştir ve miyofilamanlar boyunca
gelişigüzel yayılırlar. Sonuç olarak kalp kasında birbirinden ayrı kas lifçi-
ği demetleri bulunmaz.
Kalp kası hücrelerinde triadlara sık
olarak rastlanmaz. Çünkü T tübiilleri sadece sarkoplazma retikulıımu sarnıcının
tek yanlı genişlemesi ile birlikte izlenir. Yani kalp kası bir T tübülii ve bir
sarkoplazma retikulıımu sarnıcından oluşan diyadlar ile özellik kazanmaktadır.
Kalp kası hücreleri çok sayıda
miiokondri içerir. Bunlar sitoplazma hacminin %40’mdan fazlasını doldurur
(Şekil 10- 19). Bu durum, kalp kasının sürekli biçimde oksidatif metabolizmaya
duyduğu gereksinimi yansıtmaktadır. Karşılaştırma açısından, iskelet kasında
bu oran yalnızca %2’dir. Kalbin ana yakıtı olan yağ asitleri lipoproteinlerle
kalp kası hücrelerine taşınır. Yağ asitleri, kalp kası hücrelerinde gözlenen
çok sayıda lipit damlası içinde trigliseridler halinde depolanır. Çok az
miktarda glikojen de bulunur ve bunlar stres durumunda enerji üretimi için
glikoza yıkılabilirler. Uzun yaşayan hücreler içinde çok rastlanan Hpofuskin
pigment granül- leri (yaşlılık granülleri) kalp kası hücrelerinin çekirdek
kutuplarının yakınında bulunur.
Kalp kulakçığı kaslarıyla, kalp
karıncığı kasları arasında yapısal açıdan bazı farklar vardır. Kalp kasının her
iki tipinde de miyofilamanların düzenlenişi birbirinin aynıdır. Ancak kalp
kulakçığındaki kaslarda T tiibüllerinin sayısı dikkati çekecek kadar azdır ve
hücreler daha küçüktür. Kalp kası çekirdeğinin her iki kutbunda, her biri
yaklaşık 0.2-0.3 pm çapında, zarla sarılı ve bu bölgedeki Golgi kompleksi ile
bağlantılı granüller bulunur. Bu graniiller (Şekil 10-28) sağ kalp kulakçığındaki
kas hücrelerinde daha fazladır (yaklaşık 600/hücre), ancak sol kalp kulakçığı,
kalp karıncıkları ve vücudun başka değişik yerlerinde de bulunurlar. Kalp
kulakçığındaki bu granüller alrial natriüretik faktör olarak bilinen polipeptid bir hormonun,
molekül ağırlığı yüksek öncülünü içerir. Bu hormon sodyum ve su tutulmasına
neden olan aldesteron ve antidiüretik hormonun böbrekler üzerindeki etkilerine
karşıt yönde etki gösterir.
Kalbin zengin otonom sinir sistemi ve dizemli (ritmik)
uyartı oluşturan ve bunları ileten yapılarına 11. bölümde değinilmektedir.
Düz kaslar her biri bir bazal lamina
ve retiküler lif ağı ile kuşatılmış (Şekil 10-29), çizgilerime göstermeyen,
uzunlamasına hücrelerden oluşmaktadır (Şekil 10-30, 10-31 ve 10-32). Bazal
lamina ve retiküler lif ağı her bir düz kas lifinin oluşturduğu gücün toplu
bir harekete, örneğin, bağırsaklardaki peristaltik hareketlere dönüşmesinde rol
oynar.
Düz kas hücreleri iğ şeklindedir;
yani, en geniş yeri ortalarıdır ve uca doğru incelirler. Boyları, küçük kan
damarlarında 20 pm ile gebedeki uterus damarında 500 pm’ye dek değişebilir.
Gebelik sırasında uterus düz kas hücreleri gerek boy, gerekse sayı olarak
belirgin bir artış gösterir. İ ler bir hücre en geniş bölümü olan orta kısmında
merkezi yerleşimli tek bir çekirdeğe sahiptir. Sıkı bir şekilde bir hücrenin
ince kısmı diğer hücrenin geniş kısmıyla komşuluk yapar. Bu düzenleniş
nedeniyle enine kesitte, yalnızca geniş çaplı olan yerleri çekirdek içeren bir
dizi değişik çaplı kas hücresi görülür (Şekil 10-29). Diiz kas kasıldığı zaman
hücre kenarları katlanır ve çekirdek tirbuşon görünümü alır (Şekil 10- 33).
Mitokondri, poliribozomlar, kaba
endoplazma retikulu- mıı sisternaları ve Golgi kompleksi çekirdek kutuplarında
yoğunlaşmıştır. Hücre yüzeyine yakın bölgelerde sıkça pino- sitoz vezikiilleri
bulunur (Şekil 10-32).
Çizgili kasın sarkoplazma
retikulumuna benzer kapalı bir zar sistemi oluşturan iyi gelişmemiş bir
sarkoplazma retikulıımu bulunur. Düz kas hücrelerinde T tübülii yoktur.
Düz kasın tipik kasılma aktivitesi
aktin ve miyozin fila- manlarının yapısına ve organizasyonuna bağlıdır. Bunlar
iskelet kasında olduğu gibi açık-koyu görünüm sergilemez. Düz kas hücrelerinin
miyofilamanları hücre boyunca çapraz kesişmeler yapan, kafese benzer bir ağ
oluştururlar. Bu lıantlar 5-7 nm kalınlığında tropomiyozin ve aktin filamanla-
rından ve 12-16 nm kalınlığında miyozin fikımanlarından oluşur. Yapısal ve
biyokimyasal çalışmalar düz kaslardaki
Şekil 10-24. Kalp kasına ait iki hücrenin
uzamına kesilmiş bölümleri. İnterkalar diskin enine görülen kısımlarında
fasya adherens ve çok sayıda desmozom izlenmektedir. Uzamına izlenen bölümler
(ok) gap junctionları içerir. Bu hücrelerde çok sayıda mitokondri bulunur
(M). İki hücre arasında retiküler lifler yer almaktadır. x 18.000. (Junqueira LCU, Salles LMM:
Ultra-Estrutura e Funçao Celular. Ed-gard
Blücher’den izinle çoğaltılmıştır, 1975.)
aletin ve miyozinin çizgili
kaslardakine benzer şekilde kayan Ulaman düzeneği ile kasıldığını göstermiştir.
Diiz kas hücrelerinde kasılmanın
başlatılması için hücre içine Ca+“ girişi gerekir. Düz kastaki
miyozinin hafif zinciri foslbrillendiğinde sadece aletinle karşılıklı
etkileşimde bulunur. Hu nedenden ötürü ve troponin bulunmaması yüzünden düz
kaslarda kasılma mekanizması iskelet ve kalp kasından farklılık gösterir. Düz kaslarda
Ca2+ yine kas olmayan hücrelerin kasılmasında yer alan bir Ca2+
bağlayıcı bir protein olan kalmodülin ile birleşir. Ca-+-kalmodiilİn
kompleksi, miyozin hafif-zincir kinazmı aktive eder. Bu miyozinin
fosforilasyonundan sorumlu olan enzimdir.
Kalsiyumdan başka faktörler de
miyozinin hafif-zincir aletivitesi üzerine etkilidir ve bunlar düz kas
hücrelerinin kasılma derecesini etkiler. Kasılma ya da gevşeme döngüsel AMP
(cAMP) üzerinden hormonlarla düzenlenir. cAMP seviyeleri arttığında, miyozin
hafif-zincir kinazı aktive edilir, miyozin fosfoı illenir ve hücreler kasılır.
cAMP'delei azalma karşıt etki gösterir, kasılma azalır. Sinirsel olmayan
kontrolün baslea bir örneği seks hormonlarının uterus düz kasları üzerindeki
etkisidir. Östrojenler cAMP’yi artırır, miyozinin fosfo- rilasyonunu uyarır ve
uterus kaslarını kasar. Progesteron karşıt yönde etki gösterir. cAMP’yi
azaltır, miyozinin defos- forilasyonunu başlatır ve uterus kası gevşer.
Düz kas hücrelerinin sitoplazmasını
boydan boya kaleden K) nm’lik ara filamanlar düzenli bir dizilimine sahiptir.
Desmin (skeletin) tüm düz kaslardaki ara filamanların ana proteini olarak
tanımlanmıştır. Damar düz kaslarında ek bir

Şekil 10-25. Kalp kasının in- terkalar disk
bölgesindeki ince yapısı. Enine kesit bölgesinde hücrelerin teması girintilerle
tamamlanır; uzamına düzlemde (UD) ise temas yüzeyi düzdür. (Marshall JM. The heart, İn: Medical
Physiology,
13th ed, Vol 2, Mountcastle VB [editör]. Mosby, 1974’den izinle yeniden
basılmıştır. Fawcett DW, McNutt NS: J Celi Biol 1969;42:1 sonuçlarına göre, Poche
R Lindner E: Zellforsch Mikrosk, Anat 1955;43:104'den değiştirilerek.)
yapı da vimcntindir. Düz kaslarda iki
tip yoğun cisim gözlenir (Şekil 10-33). Birisi zara bağlı, diğeri ise
sitoplazmiktir. Her ikisi de a-aktinin içerir ve bu açıdan çizgili kasların Z
bantlarına benzer. İnce ve ara filamanların her ikisi de kasılma gücünü
bitişik düz kas hücrelerine ve bu hücrelerin çevresindeki retikiiler lif ağına
aktaran yoğun cisimlerin içine sokulurlar.
Bir düz kas demetinde sinir
dağılımının derecesi kasın işlevine ve büyüklüğüne bağlıdır. Düz kaslar otonom
sinir sisteminin sempatik ve parasempatik sinirleri ile uyarılır. İskelet
kaslarındaki nöromiisküler bağlantılar düz kaslarda bulunmaz. Çoğunlukla otonom
sinir aksonları endomisyuma ait bağ dokusunda bir dizi genişleme ile son bulur.
Genelde düz kaslar viseral boşluklu
organların, örneğin bağırsakların, uterusun ve üreterlerin duvarlarında geniş
tabakalar halinde bulunur. Hücrelerde çok sayıda aralık bağlantısı (gap
junction) vardır ve sinir desteği bakımından nispeten zayıftırlar. Bunlann
kasları toplu (sinsisyal) biçimde işlev görürler ve viseral düz kaslar
olarak
adlandırılırlar. Bunun aksine, çok birimli düz kaslar zengin bir sinir ağına sahiptir ve
böylece gözün irisinde olduğu gibi kasılmaların derecesi çok kesin bir biçimde
düzenlenebilir.


Şekil 10-27. Kalp kası uzamına kesitinin
elektron mikroskop fotoğrafı. Çizgilenme düzenine, kas lifçiklerinin değişmeli
durumuna ve kristaları bol mitokondriler dikkat çekmektedir. Fotoğrafta
kalsiyum depolama yönünde özelleşmiş düz endoplazma retikulumu olan sar-
koplazma retikulumu (SR) izlenmektedir. x 30.000.

Şekil 10-28. Çekirdek kutbunda toplanmış
natriüretik granüllerin görüldüğü kalp kulakçığındaki kas hücresinin elektron
mikroskop fotoğrafı, (izin alınarak J.C.Nogueira).

Şekil 10-29. Düz kas hücrelerinin uzamına
(üstte) ve enine (altta) kesitlerini gösteren ışık mikroskop fotoğrafları.
Çekirdeklerin merkezde olduğu görülmektedir. Çoğu hücrede çekirdekler kesite
girmez. PT boyası. Orta büyütme.
Düz kaslar genellikle sinir uyartısı
olmaksızın kendiliğinden aktive olabilir. Bu yüzden sinirleri iskelet kasında
olduğu gibi kasılmayı başlatmaktan çok düzenleme işlevi görür. Düz kaslar,
aktiviteyi uyarıcı ve baskılayıcı olmak üzere birbirinin antagonisti olarak
işlev gören hem adrenerjik vehem de kolinerjik sinir sonlanmaları alırlar. Bazı
organlarda, koli- nerjik sonlanmalar harekete geçirirken adrenerjik sinirler
baskılar; başkalarında ise tersi söz konusudur.
Kasılma aktivitesine ek olarak, düz kas hücreleri normalde
fibroblastların işleviyle ilişkili hücre dışı ürünler olan kol- lajen, elastin
ve proteoglikanları da sentezlerler.
Erişkinlerdeki 3 kas tipi hasardan
sonra yenilenme konusunda farklı özellikler gösterir.
Kalp kasının çocukluk çağının ilk
dönemlerinden sonra gerçek anlamda bir yenilenme kapasitesi yoktur. Hasar gören
ya da yıkım geçiren bölümün yerini bağ dokusunun mi- yokard nedbeleri oluşturan
bağ dokusu artışı alır.
İskelet kasında, çekirdeklerin mitoz
yeteneği bulunmamasına karşın, doku sınırlı ölçüde onarılabilir. Onarıcı hücrelerin
kaynağının uydu (satellit) hücreler olduğuna inanılmaktadır. Bunlar, her bir
olgun kas lifi etrafındaki bazal la- mina altında yer alan iğ şeklinde tek
çekirdekli hücrelerdir. Kas liflerinin yüzeyine çok yakın olduklarından
yalnızca elektron mikroskop ile görülebilirler. Bunların, kasın farklılaşmasından
sonra burada kalan inaktif miyobhıstlar oldukları kabul edilir. Zedelenmeden
ya da bazı başka uyartılardan sonra sessiz duran normal uydu hücreler harekete
geçebilir, çoğalabilir ve yeni iskelet kası liflerine dönüşebilirler. Uydu
hücrelerin benzer bir aktivitesi de yoğun egzersizden
Şekil 10-30. Bir düz kas parçasının çizimi.
Bütün hücreler retiküler liflerden oluşan bir ağ ile sarılır. Enine kesitte,
bu hücreler değişik çaplarda izlenir.
Şekil 10-31. Retiküler lifleri boyamak için
gümüşle boyanmış düz kasın enine kesiti. Retiküler lifler bu yöntemle
boyanmayan kas hücrelerinin çevresini ağ biçiminde sarmaktadır x 300.
Şekil 10-32. Düz kasın enine kesitine ait
elektron mikroskop fotoğrafı. Kesitteki hücreler değişik çaplarda görülür ve
yüzeyin hemen altında çok sayıda vezikül bulunur. Kalın ve ince filamanlar kas
lifçikleri şeklinde düzenlenmemiştir ve mitokondri (M) sayısı azdır. Retiküler
lifler ve hücreler arasında küçük miyelinsiz sinirler (N) dikkat çekmektedir.
x 6650.
|
Gevşemiş düz kas
hücresi
|
Şekil 10-33. Gevşemiş ve kasılmış durumdaki düz kas hücreleri. Hücre yüzeyinde
ve sitoplazma içinde bulunan yoğun cisimlere, sitoplazmik filamanlar tutunur.
Bu fila- manların kasılması sonucu, hücrenin boyu kısalır ve tüm kas bu şekilde
kasılmaya başlar. Kasılma sırasında hücre çekirdeğinin şekli bozulur.
sonra hücrelerin kendi ana hücreleriyle birleşerek kas
kitlesini artırma işlevidir. Bununla birlikte majör kas incinmesi ya da
dejenerasyonun ardından iskelet kasının yenilenme sığası çok sınırlıdır.
Campion DR: The musde
satellite cell: a review. Inc Rev Cytol 1984:87:225. Cantin M, Genest J: The
heart as an endocrine gland. Sci Am 1986;254:76. Cohen C: The protein switch of
muscle contraction. Sci Am 1975:233:36. Grounds MD: Age-associated changes in
the response of skeletal muscle cells to exercise and regeneration. Ann N Y Acad Sci
1998j854:78.
Düz kas etkin bir yenilenme sığasına
sahiptir. Yaralanmanın ardından tek çekirdekli canlı düz kas hücreleri ve kan
damarlarının çevresindeki perisitler (bkz. 11. Bölüm) mitoz- la çoğalarak
zedelenmiş dokunun yenilenmesini sağlar.
Yorumlar
Yorum Gönder